INTRODUCTION
Skeletal muscle exhibits a highly adaptable protein organization whose properties can be easily changed by aging, temperature, exercise, nutrition, and incretion, to name a few. Skeletal muscle adaptability results in a hypertrophied or atrophied state; these states are primarily associated with slow or fast fibers. Slow or fast fiber characteristics are attributed to changes in the population of intracellular proteins of the muscle cells [1]. This means that protein changes might control the adaptability of the skeletal muscle. One study concerning the content of αB-crystallin and tubulin in 10 days of atrophied soleus muscle was previously reported [2]. αB-crystallin and tubulin expression is dependent on muscle fiber type [3]; however, no study has investigated the response of αB-crystallin and α-tubulin with transition of myosin heavy chain (MHC) isoforms in plantar flexor muscles followed by hindlimb unloading. In this study, we focused on αB-crystallin and tubulin because these proteins exhibit functional properties associated with the adaptability of skeletal muscles.
αB-crystallin is localized in the Z-line of the sarcomere in cardiac and slow-twitch muscle [4,5]. It acts as a molecular chaperone during the folding process of other specific proteins to prevent protein degradation [1,6]. αB-crystallin is activated in a phosphorylation-dependent manner [7] and also acts to stabilize intermediate filament and cytoskeletal proteins in vitro [8,9]. Another study showed that disorganized cytoskeletal components trigger the phosphorylation of αB-crystallin through activating the p38 mitogen-activated protein kinase (MAPK) pathway [7]. αB-crystallin is also known to be upregulated in slow twitch muscles, which have high oxidative capability and higher protein turnover rates compared to fast twitch muscles, which are characterized by their high glycolytic capability [4,10].
Microtubules function as an intracellular frame to shape the cell, maintain cellular morphology, and transport proteins throughout the cell [11-13]. Microtubules exhibit a morphological, structural, and dynamic property called ‘dynamic instability’, which is regulated by the concentration of free- tubulin dimers [14,15]. Disorganized tubulin can be easily degraded through regulating this process. The degraded tubulin monomer is associated with heat shock proteins (HSPs) [16]. Microtubules are scaffolding proteins that contribute to cell adhesion [17]. Microtubules are directly associated with myosin, and the localization of myosin filaments during sarcomere formation and myogenic cell morphology is partly dependent on the microtubule networking organization [18]. It can be inferred that different muscle types are due to the different myogenic cell morphologies and their localization is related to differences in the formation and content of microtubules.
The MHC is a major component of muscle and can be transformed into various isoforms in muscle cells in response to various stimuli. The increase in directional shifts from the MHC I to the MHC IIa, MHC IId/x, and MHC IIb isoforms, augments the velocity of the shortening and power generation capacity of the MHC, but decreases its ATPase activity. This is also known as the order of MHC isoform shift and vice versa [19]. Giger et al. reported that an atrophy response due to unloading induces the shift of MHC isoforms from slow-twitch to fast-twitch muscle types [20].
It is thought that αB-crystallin, tubulin, and myosin work closely with one another during the processes of myocyte formation. αB-crystallin plays a critical role in chaeperoning for tubulin, which functions to maintain muscle cell morphology and transport organelles in the cell. αB-crystallin also indirectly supports power transmission since it acts as a chaperone protein for myosin, one of the main components of muscle. Microtubules, which are composed of tubulin, also directly associate with myosin to ensure its proper localization during sarcomere formation [5]. These data suggest that the functional relationship between αB-crystallin, tubulin, and myosin are important for muscular function and adaptability.
We hypothesized that hindlimb unloading induces muscular atrophy with MHC isoform transformation toward the MHC I→MHC IIa→MHC IId→MHC IIb isoforms. Furthermore, we evaluated whether αB-crystallin and α-tubulin would show different expression patterns in different types of atrophied muscles as atrophied slow muscle shows a more sensitive response to both proteins compared to atrophied fast muscle. We examined the triceps surae in the hindlimbs (soleus, plantaris, and gastrocnemius muscles) because these muscles might exhibit similar functions and endure similar mechanical loads compared to other muscles in the hindlimbs, such as the tibialis anterior.
We employed the unloading model to induce muscle atrophy and to quantitatively determine the relationship between the proteins (αB-crystallin and tubulin) with the transition of MHC isoforms, and its significance to the adaptability of skeletal muscles.
METHODS
Animals and unloading procedure
In this study, we utilized 12 seven-week-old adult male Wistar rats that weighed 238.2 ± 9.1 g. After 2-3 days of adaptation, the rats were randomly divided, with half placed in a control group and the half placed in an unloading group. The unloading group was tested using the hindlimb unloading model based on modified methods described by Morey and Atomi [4,21]. All animals were provided standard rat chow and water ad libitum, and were housed at 22-24 °C with a 12:12 h light-dark cycle. After 15 days, all rats were euthanized by cervical dislocation under anesthesia induced by diethyl ether for relieving pain (50 ml/kg body mass). The right sides of the whole soleus muscle, plantaris muscle, and gastrocnemius muscle were isolated, and the muscles were immediately frozen in liquid nitrogen and stored at -80 °C until use. Animal use and maintenance was approved by the University of Tokyo Animal Care and Use Committee.
Preparation of muscle samples
The three isolated whole muscles used in this study were crushed in liquid nitrogen and solubilized in a low-salt buffer containing 20 mM potassium chloride (KCL), 2 mM sodium phosphate (pH 6.8), 2 mM ethylene glycol tetraacetic acid (EGTA), 5 mM ethylene diamine tetraacetic acid (EDTA), 20 mM sodium fluoride, 1 mM sodium orthovanadate, a protease inhibitor containing 1 mM phenylmethylsulfonyl fluoride, 2 µg/ml leupeptin, 2 µg/ml aprotinin, 10 µg/ml soybean trypsin inhibitor, and a phosphatase inhibitor containing 100 nM okadaic acid and 10 mM sodium β-glycerophosphate. The homogenate was solubilized with an equal volume of 2 × sodium dodecyl sulfate (SDS) sample buffer [2 mM sodium phosphate (pH 6.8), 2% SDS, and 16% 2-mercaptoethanol] and boiled for 2 min. The homogenate was centrifuged at 15,000 × g at 4 °C for 10 min. The supernatant was collected and the protein concentration was determined using a protein determination kit (Bio-Rad; Richmond, VA, USA).
SDS polyacrylamide gel electrophoresis (PAGE) and immunoblotting
Polyacrylamide gels (8.5% to 12%) were used as running gels to detect αB-crystallin, tubulin, and myosin proteins. As stacking gels, we used 3.5% polyacrylamide gels. To separate MHC isoforms, 1.6 µg of sample was electrophoresed over a 24-h period at a constant voltage of 150 V and 4 °C [22]. Gels were stained with Coomasie Brilliant Blue R-250 dye (Sigma-Aldrich Co.; St. Louis, MO, USA). The relative percentage of MHC isoform content was calculated using imageJ (National Institutes of Health, Rockville Pike Bethesda, MD, USA). The extracted plantaris muscle was used as a reference marker for analyzing the extracted soleus muscle because not all MHC isoforms in the plantaris muscle overlap with the MHC isoforms in the soleus muscle, and vice versa. The markers used for the gastrocnemius muscle were a mixture of extracted soleus and plantaris muscles.
The following primary antibodies were used for immunoblotting: anti-α-tubulin (1:1000; Sigma-Aldrich Co.; St. Louis, MO, USA) and anti-αB-crystallin (1:5000, raised in rabbit against the C-terminal 10 peptides [(SH) KPAVTAAPKK] of human αB-crystallin). After incubation with secondary antibodies, the membrane was incubated with an enhanced chemiluminescence kit (Amersham Biosciences Corp.; Buckinghamshire, UK). αB-crystallin was expressed in Escherichia coli [23] and tubulin (phosphocellulose column-purified tubulin, PCT) was purified from the porcine brain [24].
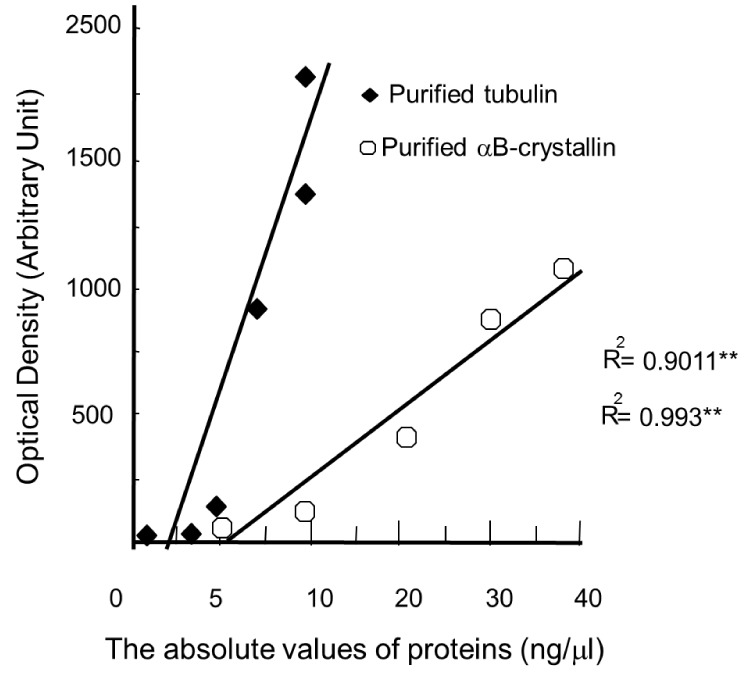
To estimate the exact quantity of each protein in the extracted samples, we used the purified proteins (αB-crystallin, 5, 10, 20, 30, and 40 ng; α-tubulin, 1, 3, 5, 7, 9, and 11 ng) that were used as standards for each protein because there was a linear relationship between the gradual contents of each protein and the band intensity (Fig. 1). We subsequently loaded different quantities of proteins extracted from the soleus (0.59 µg), plantaris (5-13 µg), or gastrocnemius (2-35 µg) muscles and their corresponding protein standards onto the same gel. The contents of αB-crystallin and α-tubulin estimated from the extracted soleus (0.5-9 µg), plantaris (5-13 µg), or gastrocnemius (2-35 µg) muscles are presented as ng/µg.
Statistical analysis
All data are presented as the mean ± standard deviation (SD) and the differences between control and experiment groups were analyzed using independent t-tests (SPSS Ver. 18.0). The statistical relation between optical density and each target protein shown in Fig. 1 was analyzed by simple regression, coefficient of determination (Fig. 1) Whole values were considered statistically significant when P < 0.05 or P < 0.01.
RESULTS
Body and muscle weight
The body and tissue weight of the control and unloading groups were measured after 15 days of hindlimb unloading (Table 1). There was a significant reduction in body weight in the unloading group compared to the control group (P < 0.05). In addition, the weights of the soleus, plantaris, and gastrocnemius muscles of the unloading groups were each significantly reduced compared to the corresponding muscle in the control group, such as 121.6 ± 18.0 (mg) to 54.9 ± 4.2 (mg), 299.3 ± 37.5 (mg) to 218.5 ± 22.9 (mg), and 1454.8 ± 140.3 (mg) to 988.3 ± 57.0 (mg), respectively (P < 0.01). Based on the whole muscle to body weight ratios, the unloading soleus (P < 0.01), plantaris (P < 0.05), and gastrocnemius (P < 0.01) muscles were significantly atrophied.
αB-crystallin content in the soleus, plantaris, and gastrocnemius muscles
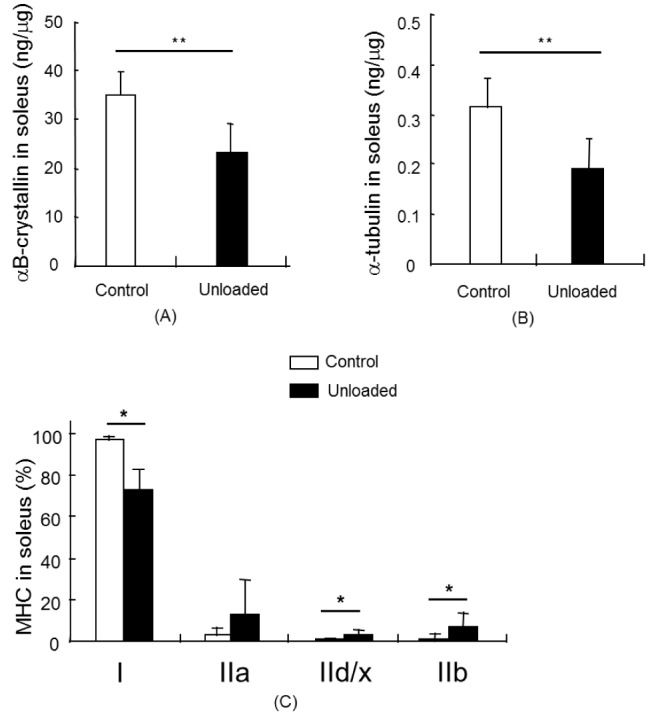
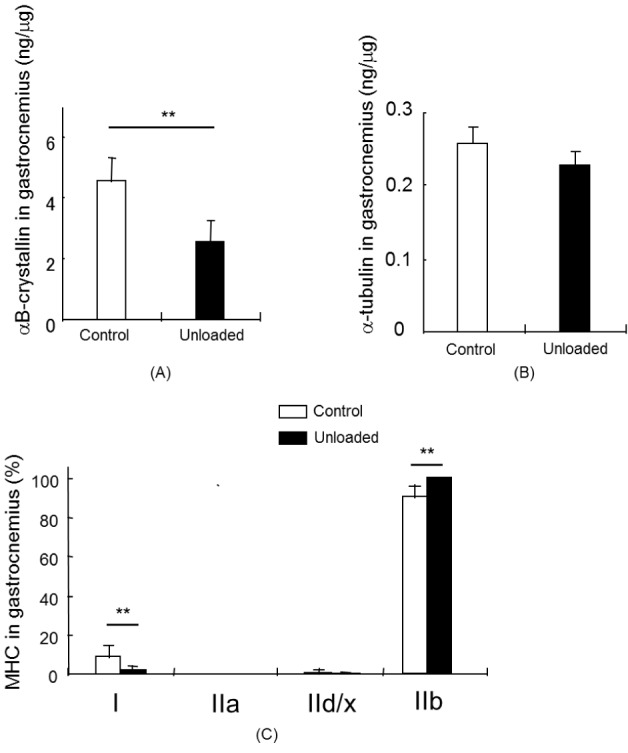
αB-crystallin levels in the soleus (P < 0.01), plantaris (P < 0.01), and gastrocnemius (P < 0.01) muscles were significantly changed in the unloading group compared to the corresponding muscles of the control group (Fig. 2a, 3a, and 4a). In the soleus muscle, αB-crystallin levels in the unloading group were approximately 28% lower than those in the control group. The αB-crystallin content in the unloading plantaris and gastrocnemius muscles was reduced by more than 37% and 42% compared to the control group, respectively. The soleus muscle contained approximately 7-fold as much αB-crystallin as the plantaris or gastrocnemius muscles did.
α-tubulin content in the soleus, plantaris, and gastrocnemius muscles
There was a significant decrease of α-tubulin in the unloading soleus muscle compared to the control muscle (P <0.01) (Fig. 2b); however, we did not observe significant differences in α-tubulin levels in the plantaris and gastrocnemius muscles between the control group and unloading group.
Percentages of the four MHC isoforms in the soleus, plantaris, and gastrocnemius muscles
The MHC isoforms in the soleus, plantaris, and gastrocnemius muscles were quantified as comparative values (Fig. 2c, 3c, and 4c). Each muscle expressed a different primary MHC isoform as shown in Figures 2c, 3c, and 4c. MHC I in the soleus muscle was significantly reduced in the unloading group compared to the control group (P < 0.05). However, MHC II was significantly increased in the unloading soleus muscle compared to the control soleus muscle (Fig. 2c). High levels of MHC IId/x and IIb expression were found in the plantaris muscle. MHC IIb was predominantly expressed in the gastrocnemius muscle. MHC IId/x was significantly reduced in the unloading plantaris muscle compared to the control plantaris muscle (P < 0.01). MHC IIa and IIb were significantly increased in the unloading plantaris muscle compared to the control plantaris muscle (P < 0.01). MHC IIb was predominantly expressed in the gastrocnemius muscle and was significantly increased after hindlimb unloading, whereas MHC I significantly decreased after hindlimb unloading (Fig. 4c) (P < 0.01).
DISCUSSION
There were obvious correlations between the changes observed in αB-crystallin, α-tubulin, and dominant MHC isoforms in the muscles. Muscle atrophy induced a reduced expression pattern of αB-crystallin and α-tubulin in plantar flexor muscles when the muscles shifted to the fast muscle fiber in the unloading group. Both proteins, αB-crystallin and α-tubulin, were downregulated in slow muscle (P < 0.01). αB-crystallin was significantly decreased in the fast muscles of the unloading group compared to the control group (P < 0.01); however, α-tubulin was not reduced.
Hindlimb unloading
Reduced total protein levels are observed when excessive catabolism overwhelms anabolism [25]. Hindlimb unloading leads to proteolytic processes related muscle atrophy. The proteolytic process induced by hindlimb unloading may be associated with changes in MHC isoforms. For example, chaperone-mediated autophagy (CMA) is believed to be related to muscle atrophy [26].
Hindlimbs are one of the most gravitationally stressed body parts in mammals. Hindlimbs are physically influenced by the gravitational pull of the earth. Hindlimb unloading leads to the loss of muscle activity in the hindlimbs against the gravitational force and induces muscular atrophy. A reduced gravitational stimulus due to suspending hindlimbs was designed to investigate the transition and degradation of proteins that contribute to hindlimb muscle atrophy. Our results are consistent with data from Sakurai et al. We show that hindlimb unloading results in reduced α-tubulin expression, followed by reduced aB-crystallin expression in atrophied triceps surae muscles [2]. Furthermore, our results show the expression pattern of these proteins in fast triceps surae muscles, plantaris, and gastrocnemius muscles, in conjunction with alterations of MHC isoforms by hindlimb unloading.
Protein transition in atrophied muscles
Skeletal muscles are categorized as slow-twitch or fast-twitch muscles based on properties related to adaptable characteristics such as, tension, contraction velocity, force, and glycolytic and oxidative metabolism [27]. Among the triceps surae muscles, we selected the soleus muscle as representative of slow-twitch muscle and the plantaris and gastrocnemius muscles as representatives of fast-twitch muscles. We investigated the responses of αB-crystallin and α-tubulin in these muscles after hindlimb unloading.
It is thought that significant changes in the protein population contribute to muscular function, generation of force, or sensitivity to gravity stimuli [28]. The different expression levels of proteins in these muscles are caused by different recruitment and loading patterns in response to mechanical unloading [29]. This suggests that the different tensile contractions of a muscle that causes different loading force also affects the quantity of tubulin [30]. Chaperone proteins are also involved in the passive and active tensile stress to protect its substrates. In this study, we showed that αB-crystallin and α-tubulin respond to unloading in MHC I-dominant slow muscle but not in MHC II-dominant muscles.
In our study, the α-tubulin and αB-crystallin levels exhibited similar changes between control and unloading soleus muscle (Fig. 2b). However, we did not observe significant changes in α-tubulin levels between the control and unloading group in the plantaris and gastrocnemius muscles (Fig. 3b, and 4b).
Sakurai et al. reported that the β-tubulin transcript level did not change after 10 d of tail suspension even though the αB-crystallin transcript level was attenuated when compared to controls [2]. Based on a previous report [2] and our results, tubulin appears to have a less sensitive response than does αB-crystallin. In atrophied muscles, tubulin begins to degrade after αB-crystallin has degraded. Tubulin is a scaffold protein, and is a net component of the cytoskeleton that maintains the cellular structure [2,31]. The hypothesis that the degradation of tubulin accelerates muscular atrophy is supported by the fact that the most significant decrease in tubulin levels (P < 0.01) was observed in the atrophied soleus muscle, which also showed the greatest extent of atrophy among the three skeletal muscles (47% reduction). By contrast, relatively less prominent atrophy was observed in the gastrocnemius (21% atrophied) and plantaris (16% atrophied) muscles, in which tubulin was not significantly altered when compared to the controls.
Role of MHC in atrophied muscles
MHC is also associated with αB-crystallin [7,32]. αB-crystallin assists in preventing the aggregation of myosin and maintains ATPase activation [32]. The hydrophobic patch of αB-crystallin regulates unfolded domains of myosin heavy chain [32]. It is thought that hindlimb unloading directly or indirectly affects the activation of quiescent satellite cells in the disruption of sarcolemma. These stimulated satellite cells activate hepatocyte growth factor (HGF) and the activated HGF initiates a signaling cascade, such as the 38p MAPK pathway [7]. This signaling pathway is likely related to the expression of aB-crystallin and the MHC isoforms in this study. We used 4 MHC isoforms to reliably identify the dominant MHC isoform in slow-twitch and fast-twitch muscles [33]. We utilized these isoforms because the dominant MHC isoform is partly related to the morphological, contractile, metabolic, and fatigue-resistant properties of muscles [34-36]. We analyzed whole plantar flexor muscles because a whole muscle unit, such as the soleus, plantaris, and gastrocnemius muscles, can be linked to its explicit function and adaptability, which were the factors of interest for this study, and because at the molecular level, the border of the medial or lateral portion of the muscle is quite ambiguous, which could lead to inconclusive or erroneous results [37]. Our results indicate that the atrophied plantar flexor muscles began to shift and resemble a fast twitch muscle. This is consistent with other studies that demonstrated a fiber type transition caused by atrophy mainly due to a reduction in the fiber size of the muscle [20]. These results may be related to αB-crystallin and tubulin expressions since the expressions of these proteins are dependent on fiber type [3]. We could not clearly detect changes of tubulin because we used whole fast muscles in this study; therefore, in future studies, we will analyze changes of tubulin in muscle fibers. In addition, future studies will also focus on identifying changes in protein levels in hypertrophied slow and fast muscles.
CONCLUSION
In conclusion, we found that αB-crystallin and tubulin related to the transition of MHC isoforms were downregulated more explicitly in the atrophied soleus muscle rather than in the atrophied plantaris and gastrocnemius muscles. This finding may explain the phenomenon that the target proteins in the fast muscles reacted less sensitive to microgravity than those in the slow muscle. Further studies are needed to provide more clear mechanism regarding the adaptability of skeletal muscles from disuse or immobilization.