



INTRODUCTION
Advanced glycation end products (AGEs) are the non-enzymati-cally modified forms of proteins or lipids generated upon exposure to sugars. AGEs form in vivo in hyperglycemic environments and during aging, and contribute to the pathophysiology of vascular complications in diabetes1, 2.
Glycation is a spontaneous non-enzymatic reaction of free-reducing sugars with the free amino groups of proteins, DNA, and lipids, resulting in the formation of an Amadori product. The Amadori product irreversibly undergoes a variety of dehydration and rearrangement reactions, leading to the formation of AGEs. AGEs accumulate in vascular wall tissues and on plasma lipoproteins, and bind to AGE-specific receptors (RAGEs); AGEs bind to RAGE at an accelerated rate in diabetic patients, and play an important role in the development of diabetic complications3. Moreover, AGEs can covalently cross-link and biochemically modify protein structures, and affect the function of proteins, particularly collagen. The diabetes-induced modification of long-lived proteins, such as collagen and lens crystallin by glycation is represented an increase in the fluorescence and cross-linking of the protein4. The irreversible formation of AGEs and their cross-linking with proteins cause damage to the kidneys, eyes, and blood vessels5, 6.
Aminoguanidine was first introduced as a AGE inhibitor7. As described in previous reports, aminoguanidine prevents the renal, retinal, and neural complications of diabetes through the inhibition of AGE formation8. However, owing to the safety concerns due to its adverse effects including pro-oxidant activities9 and inhibition of NO synthase10, aminoguanidine cannot be used clinically11.
Since herbal products have generally been proven to be safer for human consumption compared with synthetic compounds, there has been an increasing interest in the use of botanical compounds as anti-AGE agents12. Panax ginseng is a widely used herbal supplement, and has been used traditionally for centuries in Asian countries to promote vitality. Red ginseng, which is the processed root of P. ginseng product, is manufactured through cycles of steaming and drying13. This manufacturing process results in the formation of additional beneficial compounds known as ginsenosides. Red ginseng has shown potent pharmacological activities against immune responses, metabolic diseases, and cancer14-16. Recently, some methods of transformation including enzymatic conversion17 and fermentation18 of ginsenosides from red ginseng have been used. These biotransformation processes of ginsenosides from red ginseng have increased its pharmacological potency in several animal disease models19-22. GS-E3D is a newly developed pectin lyase-modified red ginseng extract. This product has been shown to exhibit anti-obesity effects in a mouse model23, and anti-inflammatory activities in macrophage cells in vitro24. To the best of our knowledge, there have been no studies on the anti-glycation activity of GS-E3D. To address this issue, we studied the efficacy of GS-E3D as an AGE inhibitor in vitro and in vivo. In this study, the effectiveness of GS-E3D was compared with that of the well-known AGE inhibitor, aminoguanidine.
METHODS
GS-E3D preparation
The material used in this experiment was a 4-yearolddried P. ginseng root purchased from a local market(Wooshin Industrial Co., Ltd., Geumsan, Korea), andwas deposited in the International Ginseng and HerbResearch Institute (No. GS201104). GS-E3D was preparedaccording to our previous report23. Briefly, redginseng extract, which was adjusted to 6┬░ Brix wasincubated with 10% pectin lyase (EC 4.2.2.10, Novozyme, #33095, Bagsvaerd, Denmark) at 50┬░C for 5 daysin a shaking incubator (150 rpm). To terminate the reaction, the processed extracts were heated at 95┬░C for 10min, and then freeze-dried. The dried GS-E3D consistedof 120.2 mg/g crude saponin containing the followingginsenosides: 5.9 mg/g Rg1, 12.6 mg/g Re, 4.7 mg/gRf, 30.2 mg/g Rb1, 14.0 mg/g Rc, 17.6 mg/g Rb2, 2.5mg/g Rb3, 27.7 mg/g Rd, 1.3 mg/g 20(S)-Rg3, 1.4 mg/g20(R)-Rg3, 0.8 mg/g Rk1, and 1.5 mg/g Rg5.
Inhibitory effect on AGE formation in vitro
AGEs were produced in the in vitro system by a mo-dified method that has been previously described25. Bovine serum albumin (10 mg/mL, Sigma Chemicals, MO, USA) was incubated at 4┬░C for 7 days with methylglyoxal (5 mM) in sodium phosphate buffer (0.1 M, pH 7.4). All of the reagents and samples were sterilized by filtration through 0.2 mm membrane filters. This reaction mixture was then mixed with GS-E3D. Aminoguanidine (Sigma Chemicals, MO, USA) was used as a positive inhibitor. The fluorescence intensity of fluorescent AGE formation was measured using a spectrofluorometric detector (BIO-TEK, Synergy HT, Ex: 350 nm, Em: 450 nm). The concentration of each test sample resulting in 50% inhibition of the activities (IC50) was estimated from the least squares regression line of the logarithmic plot of concentration against the remaining activity.
Inhibitory effect on AGE cross-linking
The ability of compounds to inhibit AGE cross-linking was measured by a previously reported method26. Briefly, the mixture of 1 ╬╝g AGE modified bovine serum albumin (BSA) (Cosmo Bio, Tokyo, Japan) with either test concentrations of GS-E3D or aminoguanidine was added to each well of collagen-coated microtiter plates (Sigma, MO, USA). AGE-BSA was allowed to react with collagen for 4 h at 37┬░C. The formation of the collagen-AGE-BSA complex was detected using an anti-AGE monoclonal antibody (6D12, Cosmo Bio, Tokyo, Japan), a horseradish peroxidase-conjugated goat anti-mouse IgG antibody, and a H2O2 substrate containing ABTS chromogen. The optical density (OD) at 410 nm was measured on an ELISA reader (BIO-TEK, synergy HT). The inhibition of cross-linking was expressed as the percentage decrease in OD when AGEBSA was incubated with collagen in the presence of the compounds.
Cross-link breaking effect on preformed AGE cross-links
The ability of GS-E3D to break preformed AGEs was measured by a previously reported method with minor modifications27. Briefly, 1 ╬╝g of glycated bovine serum albumin (AGE-BSA, MBL International, MA, USA) was preincubated in collagen-coated 96-well plates (Nunc, Roskilde, Denmark) for 24 h, and the collagen-AGE-BSA complexes were incubated with or without GS-E3D (Sigma, MO, USA) or alagebrium (Suchem Pharma Co., Wenzhou, China). The collagen-AGE-BSA cross-linking was detected using an mouse anti-AGE antibody (6D12, Wako, Osaka, Japan), a horseradish peroxidase-linked anti-mouse IgG antibody, and a substrate containing 3,3ŌĆ▓,5,5ŌĆ▓-tetramethylbenzidine chromogen. The levels of cross-link breakage were calculated as the percentage decrease in optical density at 410 nm. We calculated the inhibitory concentration 50% (IC50, ╬╝g/mL) as 50% inhibition of the collagen-AGE-BSA cross-linking.
Animals
Seven-week-old male Sprague-Dawley rats were purchased from Orient Bio (Seongnam, Korea), and acclimated for 1 week prior to the study. Diabetes was induced by a single injection of streptozotocin (STZ, 60 mg/kg, i.p.). The age-matched control rats received an injection of an equal volume of vehicle (0.01 M citrate buffer, pH 4.5). One week after the STZ injection, a blood sample was obtained from the tail vein. Rats with a blood glucose level over 300 mg/dL were defined as diabetic rats. The rats were randomly divided into 5 groups of 10 each as follows: (1) normal control rats (NOR), (2) STZ-induced diabetic rats (DM), and (3, 4, and 5) STZ-induced diabetic rats treated with GS-E3D (25, 50, and 100 mg/kg body weight, respectively). GS-E3D was dissolved in the vehicle (distilled water). GS-E3D was orally administered to the rats for 6 weeks. All experimental procedures were performed under the supervision of our Institutional Animal Care and Use Committee (IACUC approval No. 15-100).
Quantification of AGE formation in vivo.
To determine AGE formation, serum samples were analyzed by a competitive enzyme-linked immu-nosorbent assay (ELISA). The assay was performed using a monoclonal AGE antibody (6D12, Cosmo Bio, Tokyo, Japan) according to established protocols28.
RBC-IgG assay
Immunoglobulin G (IgG) is observed to be cross-linked to the membrane proteins of red blood cells (RBCs). RBC-IgG complexes are formed before other AGE cross-links in vivo. The amount of RBC-IgG can be used to estimate the levels of protein cross-linking27. To test the inhibitory effect of GS-E3D on AGE cross-linking, RBCs from heparinized whole blood were collected, and RBC-IgG levels were determined using anti-IgG ELISA.
RESULTS
Inhibitory effect of GS-E3D on AGE formation in vitro
GS-E3D was analyzed by in vitro bioassays to eva-luate AGE-BSA formation. The inhibitory effect of GS-E3D on AGE-BSA formation is summarized in Table 1. GS-E3D inhibited the formation of AGE-BSA (IC50 = 19.65 ┬▒ 4.35 ╬╝g/mL). The inhibitory activity of GS-E3D was stronger than that of aminoguanidine (IC50 = 80.28 ┬▒ 3.39 ╬╝g/mL) and the unmodified red ginseng extract (IC50 = 139.46 ┬▒ 68.18 ╬╝g/mL).
Inhibitory effect of GS-E3D on the cross-linking of AGEs with collagen in vitro
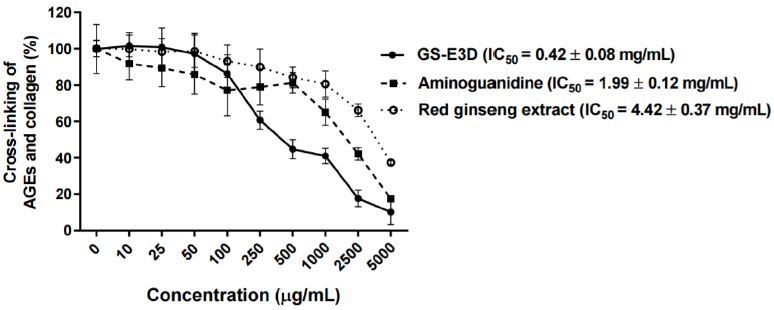
The inhibition of the cross-linking of AGE-BSA withcollagen at various concentrations of GS-E3D was tested(Figure 1). GS-E3D decreased the cross-linking of AGE-BSA with collagen in a dose-dependent manner; the IC50value of GS-E3D was 0.42 ┬▒ 0.08 mg/mL, and its inhibitoryactivity was stronger than that of aminoguanidine (IC50 valueof 1.99 ┬▒ 0.12 mg/mL) and the unmodified red ginsengextract (IC50 = 4.42 ┬▒ 0.37 ╬╝g/mL).
Figure┬Ā1.
Inhibitory effect of GS-E3D on the cross-linking of AGEs with collagen in vitro. The cross-linking of AGE-BSA with collagen was detected by ELISA. Data are presented as means ┬▒ SE (n = 4). The IC50 value was calculated from the dose-inhibition curve. Aminoguanidine was used as the positive control.
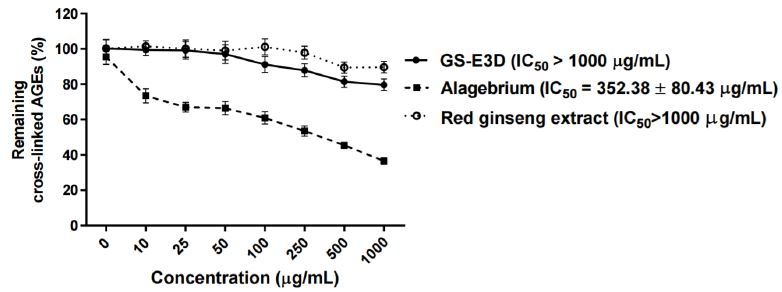
Effect of cross-link-breaking of GS-E3D on preformed AGE cross-links with collagen in vitro
We tested whether GS-E3D could also interact with preformed AGEs in vitro. As shown in Figure 2, incubation with GS-E3D, the unmodified red ginseng extract or alagebrium over a range of concentrations destroyed the preformed AGE-BSA-collagen cross-links. Alagebrium, which is a well-known AGE-breaker, dose-dependently destroyed the cross-links in the preformed AGE-BSA complexes with rat-tail tendon collagen (IC50 = 352.38 ┬▒ 80.43 ╬╝g/mL). However, GS-E3D and the unmodified red ginseng extract did not break the cross-links of AGE with collagen.
Figure┬Ā2.
Effect of cross-link-breaking of GS-E3D on the preformed cross-links of AGEs with collagen in vitro. The cross-linking of AGE-BSA with collagen was detected by ELISA. Data are presented as means ┬▒ SE (n = 4). The IC50 value was calculated from the dose-inhibition curve. Alagebrium was used as the positive control.
Blood glucose
Blood glucose levels are summarized in Table 2. Blood glucose levels had significantly increased in the diabetic rats (p < 0.05). No differences in blood glucose levels were observed between the GS-E3D-treated and vehicle-treated diabetic rats.
GS-E3D inhibits AGE formation in vivo and AGE cross-linking
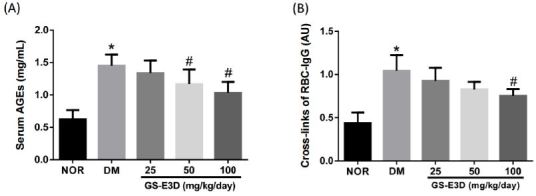
The ability of GS-E3D to inhibit AGE formation in vivo was tested. At the end of the study, the AGE levels in serum were remarkably elevated in the vehicle-treated diabetic rats compared to the normal control rats. However, these levels in the GS-E3D-treated diabetic rats had considerably decreased compared to the vehicle-treated diabetic rats (Figure 3A). Next, we carried out an RBC-IgG assay to evaluate AGE cross-linking. As shown in Figure 3B, the RBC-IgG level in the vehicle-treated diabetic rats had substantially increased compared to that in the normal control rats. However, treatment with GS-E3D considerably reduced the level of RBC-IgG compared to the vehicle-treated diabetic rats.
Figure┬Ā3.
Effects of the in vivo treatment with GS-E3D on AGE formation (A) and IgG cross-linking with the RBC surface (B) in the blood of the streptozotocin-induced diabetic rats. The values in the graph represent means ┬▒ SE (n = 10). *p < 0.05 vs. normal control rats, #p < 0.05 vs. vehicle-treated.
DISCUSSION
Many dietary supplements are being sold with advertisements of their numerous beneficial effects. GS-E3D is a commercial pectin lyase-modified red ginseng extract containing a high level of the ginsenoside Rd. In the present study, we demonstrated that the newly developed pectin lyase-modified red ginseng extract, GS-E3D had an inhibitory effect on AGE formation and the cross-linking of AGEs with collagen in vitro and in vivo.
It is well established that AGE formation plays a crucial role in the development of diabetic complications29. The AGE cross-links are permanent, and irreversible complexes are formed when glucose binds to the target protein, such as collagen. The cytotoxic roles of AGEs in diabetes have been shown in a number of studies30. AGEs can accumulate in many tissues of patients with diabetes. Since the body does not contain any enzyme capable of structurally degrading the AGEs, they accumulate during the biological life of the proteins31.
There is considerable interest in agents that inhibit the formation of AGEs and their cross-links or those that can break the AGE cross-links due to their therapeutic potential6, 32. Several synthetic and natural agents have been proposed as AGE inhibitors. Reactive carbonyl species are potent precursors in the formation of AGEs and cross-linking of proteins33-36. AGE inhibitors, including aminoguanidine and pyridoxamine prevent AGE accumulation by interacting with the reactive carbonyl species and acting as carbonyl traps37, 38. However, owing to safety concerns, aminoguanidine is not currently used11. Recently, several researchers have suggested that a novel agent can destroy preformed AGE-derived protein cross-links. The first AGE breaker to be identified, PTB was introduced in 1996. Since PTB is unstable in vitro, it was not clinically successful. Another compound, alagebrium39 was developed as an AGE breaker. Alagebrium could reverse AGE accumulation in vivo40. Since the clinical studies on these compounds were terminated, none of the known AGE breakers is in clinical use.
Our previous studies showed that some natural herbal products have potent anti-AGE activities41, 42. Quan et al. reported that Korean red ginseng reduced the formation and secretion of AGEs in the kidneys of diabetic rats43. The ginsenoside Rd is one of the bioactive compounds present in red ginseng, and it ameliorated the damage to astrocytes induced by methylglyoxal, which is a precursor of AGEs44. Since GS-E3D has an high level of Rd compared with an unmodified red ginseng extract45, GS-E3D may exert a more potent inhibitory effect than the unmodified red ginseng extract on the formation of AGEs and their cross-linking with proteins. Although GS-E3D has potent inhibitory effects on AGE formation in vitro and in vivo, the mechanism of its action is still not clear. Based on our findings, the inhibition of the formation of AGEs and their cross-links with proteins by GS-E3D might ameliorate the AGE burden in the diabetic rats. Furthermore, these data support the premise that GS-E3D is effective for the treatment of AGE-related diabetic complications due to the inhibition of AGE formation in various tissues and in the serum.
In conclusion, our study showed that GS-E3D is a potent inhibitor of the formation of AGEs and their cross-linking with proteins. Although we did not compare the effects of GS-E3D with those of an unmodified red ginseng extract in the animal model, GS-E3D has more potent anti-AGE activity than the unmodified red ginseng extract in vitro. Therefore, GS-E3D could be a promising drug candidate for the treatment of AGE-related diseases by reducing AGE burden.




