INTRODUCTION
Exercise-induced reactive oxygen species (ROS) act as signaling factors for physiological adaptations and pro-inflammatory responses. Exercise patterns, such as intensity and duration, affect ROS generation [1]. Short-duration exercise intermittently produces low to moderate levels of ROS, activates signaling pathways, and promotes cellular adaptations, all of which exert a protective effect against subsequent stress [2]. Conversely, high levels of ROS induced by prolonged or high-intensity exercise may cause structural and functional damage [3]. In particular, excessive ROS production during strenuous exercise impairs physiological functions, such as blood flow-mediated vasodilation and muscle contraction [4-6]. Reid [4] showed that redox interventions are important factors in exercise performance. ROS are generated during high-intensity exercise as a consequence of increased aerobic metabolism [7-9]. Excessive ROS may cause oxidative damage to tissues and cells [10] and decrease muscle function [5,6]. Highly trained athletes perform high-intensity exercises and are exposed to high levels of ROS, which may lead to physical dysfunction and a decline in performance [4,11]. In addition to the high levels of ROS, antioxidant defense systems, such as enzymes (superoxide dismutase, catalase, glutathione peroxidase, etc.) and exogenous antioxidants (vitamins, polyphenols, etc.) are activated. Therefore, it would be beneficial for athletes to take adequate amounts of antioxidants during high-intensity exercise to enhance the capacity of exogenous antioxidants.
However, there is inconsistent evidence on whether antioxidant supplementation improves athletic performance [12]. Previous studies have shown that long-term (10 to 11 weeks) high-dose vitamin C and E intervention negatively [13] or ineffectively [14] affects muscle performance and may induce maximal voluntary contraction. Although vitamin C effectively suppressed inflammation and muscle damage via short-term intervention [15], a recent systematic review confirmed that vitamins C and E did not have a performance-modulating effect [12,16].
In contrast, low to moderate doses of food-derived polyphenols improve muscular function in long-term (10 weeks) [17] and short-term (7 to 8 days) interventions through their antioxidant capacity [18-22]. These results suggest that the effect of antioxidants on exercise adaptation depends on the type and dose of antioxidants administered. In particular, an appropriate amount of food-derived polyphenol supplementation may benefit muscle performance, even with short-term loading [23]. However, negative or ineffective responses that interfere with exercise adaptation may occur due to the administration of wrong dosages or durations and low bioavailability of antioxidants [24,25].
Generally, polyphenols have low in vivo bioavailability due to several factors, such as their interaction with the food matrix during digestion and metabolic processes mediated by the liver, intestine, and microbiota [26]. However, oligomerized lychee fruit extract (OLFE) is manufactured as a low-molecular-weight polyphenol and has higher bioavailability and stronger antioxidant capacity because it is hydrolyzed and stabilized at a smaller molecular size through manufacturing. Indeed, serum polyphenol levels increase upon in vivo intervention with OLFE supplementation compared to lychee fruit intake [27]. In addition, OLFE suppresses ROS production and inflammation in humans and animals [28-30]. These properties enable safe and effective low-dose antioxidant supplementation, which may contribute to performance improvement in various exercise-related scenarios.
In the athletic field, high-intensity exercise-induced inflammation is significantly reduced after a 100 mg/day OLFE intervention for two months in male endurance athletes31 and four weeks in healthy young men [28]. Tsukamoto et al. showed that supplementation with OLFE at a 200 mg/day dose for two weeks elevates the maximum oxygen uptake in 20 sets of high-intensity intermittent exercise (HIIE) in male athletes [32]. Another study showed that supplementation with the same dose for 26 days decreased subjective fatigue in athletes after training [33]. These results suggest that OLFE supplementation from 100 to 200 mg/day enhances aerobic capacity and reduces exercise-induced inflammation without harmful effects.
HIIE requires both aerobic and anaerobic capacities. It is applied in various sports, repeating high-intensity exercise intermittently with short-term recovery or low-intensity exercise in between [34]. At the beginning of HIIE, anaerobic energy metabolism, such as ATP-PCr and the glycolysis cycle, dominates aerobic energy metabolism. In contrast, aerobic energy metabolism dominates as exercise duration increases [35,36].
In this study, we hypothesized that supplementation with OLFE may improve HIIE performance after seven days. Therefore, we conducted a pilot study to evaluate the safety and efficacy of OLFE intake on exercise performance in athletes.
METHODS
Participants
Ten male athletes from a university sports club specializing in intermittent exercise (age: 21.5 ± 0.8 y, height: 171.7 ± 4.1 cm, weight: 69.2 ± 6.4 kg) were recruited. The ethics committee of Nippon Sport Science University (No. 019-H045) and the University Hospital Medical Information Network Clinical Trials Registry in Japan (No. UMIN000041049) approved this study. After being informed of the purpose of the study, methods, possible health hazards, risks, privacy protection, data management, and publication, all participants signed a consent form. None of the participants had an ongoing or prior chronic disease or a history of smoking before enrolment in this study.
All participants were instructed to avoid using supplements and medicines that could affect OLFE supplementation during the intervention period. The body composition of the participants was measured using bioelectrical impedance analysis (InBody 730, InBody Co., Ltd., Seoul, Korea), and their physical characteristics are shown in Table 1. Participants trained five to six times a week, and their average maximum power output for the Wingate test at the baseline was 10.9 ± 0.9 W/kg. As 9 out of 10 participants competed in the same sports club and had similar training regimes, there was no significant difference in the training load between participants and phases during the intervention period. In addition, there were no official matches or other athletic events that took place during the intervention period.
Experimental design
This study was designed as an open-label trial consisting of control intervention (CON) and OLFE intervention (OLI) phases. All participants practiced before the trial to confirm the experimental protocol. A washout period of 7 days was set between the CON and OLI phases. All participants refrained from high-intensity exercise and drinking alcohol one day before the trial. They had the same meals at the same time and avoided caffeine and alcohol on the trial day. Body composition and subjective fatigue were assessed pre-trial for the CON and OLI phases. The degree of subjective fatigue was measured before exercise (at rest) using a visual analog scale method, and the participants were asked to indicate the degree on a 100 mm horizontal line.
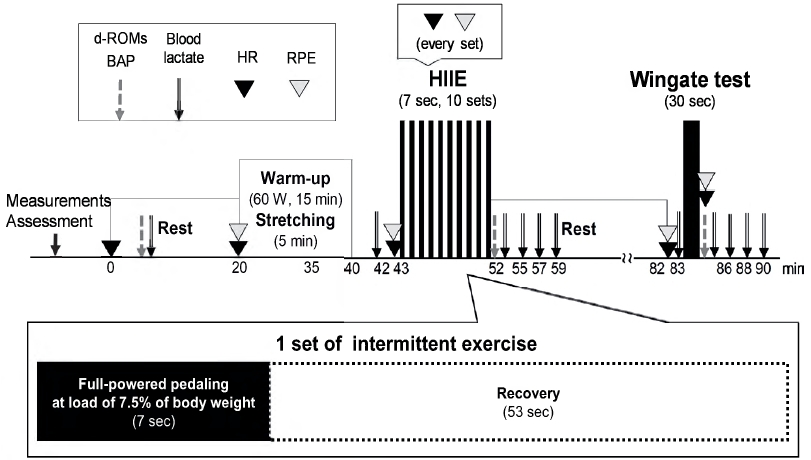
The participants performed a warm-up exercise for 15 min with a load of 1.0 kpc at 60 rpm using a bicycle ergometer (Fujin Raijin, Ohashi Chiso Laboratory Co., Ltd., Japan). After sitting for 20 min, HIIE was performed at a load of 7.5% of the bodyweight for 10 sets. The subjects rested for 30 min, and then a Wingate test was performed for 30 s under the same load. Power output, blood lactate levels, levels of reactive oxygen metabolites (diacron-reactive oxygen metabolites [d-ROMs]), biological antioxidant potential (BAP), heart rate, and rate of perceived exertion (RPE) were measured before, during, and after exercise (Fig. 1). The participants were allowed to drink water during the trial freely.
Supplementation
Oligonol (Amino Up Chemical Co., Ltd., Sapporo, Japan) supplement extracted from lychee fruit pericarp, which was mechanically oligomerized to polyphenol polymers, was used. It contains low-molecular-weight oligomerized polyphenols (100 mg per capsule). In addition, 15.7 w/w % polyphenol monomers ((+)-catechin and (-)-epicatechin) and 13.3 w/w % polyphenol dimers (procyanidin B2) were also mixed to stabilize the capsule. All participants were administered with two capsules of OLFE per day, after breakfast and dinner, for seven days in the OLI phase. The total daily intake of polyphenols was 200 mg, as reported previously [33].
Power output measurement
During the HIIE and the 30-s Wingate test, power output was calculated every 0.1 s using a bicycle ergometer. One HIIE set consisted of full-powered pedaling for 7 s at a load of 7.5% body weight, followed by 53 s of recovery. The exercise was repeated in ten sets. The average power output was calculated for each set and corrected according to body weight. The Wingate test was performed for 30 s with a load of 7.5% of the body weight. The average power output, maximum power output, and rate of power decline over a 30-s period were calculated and corrected according to body weight.
Blood biomarker measurements
Briefly, 100 μL of blood was collected from the fingertip to measure blood lactate, d-ROMs, and BAP. Blood lactate levels were measured using a lactate analyzer (Lactate Pro2, ARKRAY, Inc., Kyoto, Japan) at rest, before exercise, immediately after exercise, and at 3, 5, and 7 min after exercise. The highest value of blood lactate immediately and at 3, 5, and 7 min after exercise was selected as the blood lactate level (mmol/L) after exercise. Blood samples were collected in heparinized vacuum tubes (Microvette, SARSTEDT AG & Co. KG, Nümbrecht, Germany) and centrifuged at 2,000 × g at room temperature for 2 min.
Plasma samples were used to measure d-ROMs and BAP using a free radical analyzer (FREE Carrio Duo, Diacron International s.r.l., Grosseto, Italy) [37,38] at rest and immediately after each exercise.
For d-ROM (U.CARR) measurements, 20 μL of plasma was added to a mixture containing an acidic buffer solution (pH 4.8) and an aqueous aromatic amine solution. A chromogen was added to the mixed sample and transferred to a spectrophotometer. The plasma hydroperoxide concentration was calculated from the percentage absorbance change at 505 nm during 5 min of incubation, which estimated the d-ROM (U.CARR) levels.
BAP (μmol/L) was measured via the change in the concentration of ferric ions (Fe3+) to ferrous ions (Fe2+) reduced by the endogenous antioxidants and the exogenous antioxidants in the blood [39]. Briefly, 10 μL of plasma was added to the reaction mixture, and the absorbance was measured at 505 nm. The sample was incubated at 37 °C for 5 min, and the absorbance at 505 nm was measured again. The percentage change in absorbance was calculated from the oxidized iron ion concentration, which provided an estimated BAP.
Heart rate and RPE
Heart rate (bpm) was measured every second using a heart rate sensor (H10, POLAR M200, POLAR, Polar Electro, Kempele, Finland). Maximum values were set at each minute of HIIE and 30 s of full-powered exercise. The RPE consisted of 15 levels (6-20) of the Borg scale and was measured immediately after warming up, before, during, and after each exercise.
Statistical analysis
All data are presented as the mean ± standard deviation (SD). Differences between the CON and OLI phases were compared using the paired t-test or two-way repeated-measures analysis of variance (ANOVA). When significant differences were determined using ANOVA, post hoc analyses were conducted using the Bonferroni test. Nonparametric statistical analysis was performed for parameters without normality. Cohen’s d was calculated to measure the effect size between the control and intervention groups. Statistical analysis was performed using SPSS ver.25 (IBM Japan Inc., Tokyo, Japan), and a P-value < 0.05 was considered statistically significant.
RESULTS
Pre-trial findings
We found no significant differences in body weight (kg), body mass index, body fat (%), perception of fatigue (mm) (CON: 75.6 ± 6.9, OLI: 74.0 ± 12.8), and sleep duration (hr) (CON: 6.9 ± 1.7, OLI: 6.7 ± 1.4) between the CON and OLI phases. The average room temperature (°C) (CON: 23.7 ± 1.0, OLI: 23.6 ± 0.8) and humidity (%) (CON: 36.1 ± 5.8, OLI: 36.8 ± 5.6) were not different.
Power output
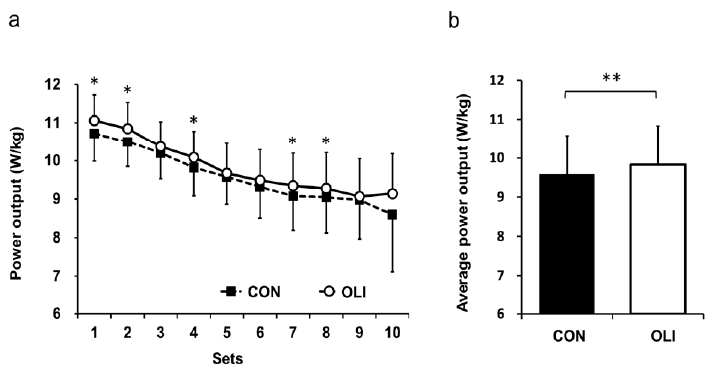
The average power output (W/kg) of sets 1, 2, 4, 7, and 8 was significantly higher in the OLI phase than that in the CON phase (P < 0.05) (Fig. 2a), and the average power output (W/kg) of ten sets in the OLI phase (9.8 ± 0.6) was significantly higher than that in the CON phase (9.6 ± 0.6) during HIIE (P < 0.01, d = 0.40) (Fig. 2b).
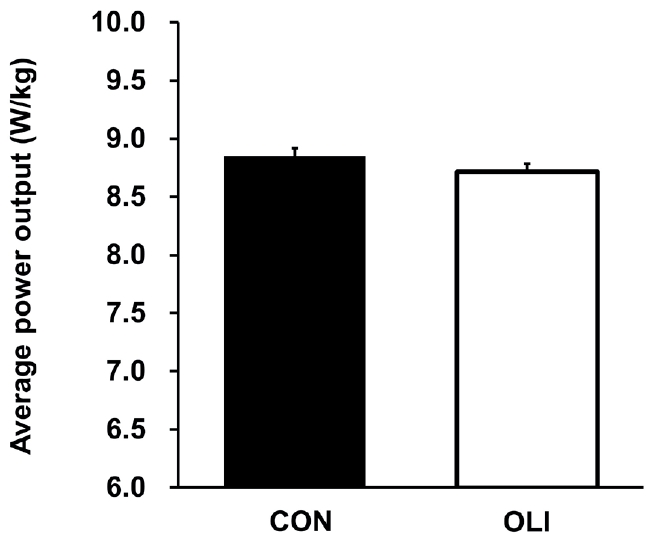
In addition, the average power output (W/kg) of the 30-s Wingate test did not differ between the CON (8.9 ± 0.8) and OLI (8.7 ± 0.7) phases (Fig. 3). Furthermore, the maximum power output (W/kg) (CON: 11.2 ± 1.1, OLI: 11.3 ± 0.9) and the rate of power decline (%) (CON: 45.9 ± 13.2, OLI: 47.1 ± 12.6) during the 30-s Wingate test were also not different between the CON and OLI phases.
Blood lactate levels
The blood lactate level (mmol/L) was not significantly different between CON and OLI phases before (CON: 1.1 ± 0.1, OLI: 1.2 ± 0.4) or after (CON: 13.8 ± 2.4, OLI: 12.7 ± 2.5) HIIE. However, the change (%) was significantly lower in the OLI phase (1078.9 ± 258.1) than that in the CON phase (1270.6 ± 291.0) in HIIE (P < 0.05, d = 0.70) (Fig. 4).
Similarly, the blood lactate level (mmol/L) was not significantly different between the CON and OLI phases before (CON: 6.3 ±2.3, OLI: 6.1 ± 1.6) and after (CON: 14.9 ± 2.4, OLI: 14.3 ± 2.0) the Wingate test, and the change (%) was not different between the CON (266.4 ± 85.7) and OLI (245.8 ± 47.2) phases.
ROS production and antioxidant capacity
The level of d-ROMs (U.CARR) did not differ between the CON and OLI phases before (CON: 286.3 ± 47.3, OLI: 283.9 ± 47.2) and immediately after (CON: 313.5 ± 55.8, OLI: 310.0 ± 53.0) HIIE, and there was no difference in the change (%) between the CON (109.4 ± 41.8) and OLI (109.5 ± 9.1) phases (Fig. 5a). Also, d-ROM (U.CARR) was not significantly different between the CON and OLI phases before (CON: 286.3 ± 47.3, OLI: 283.9 ± 47.2) and after (CON: 301.7 ± 40.8, OLI: 299.1 ± 55.2) the Wingate test, and the change (%) was not different between phases (CON, 106.0 ± 4.7; OLI, 105.3 ± 5.9).
Regarding BAP (μmol/L), there was no significant difference between the CON and OLI phases before (CON: 2085.9 ± 143.6, OLI: 2006.2 ± 205.4) and immediately after (CON: 2658.3 ± 241.9, OLI: 2698.5 ± 180.1) HIIE. However, the change in BAP (%) tended to be higher in the OLI phase (135.3 ± 10.0) than in the CON phase (127.4 ± 6.2) during HIIE (P = 0.06, d =1.22) (Fig. 5b). Also, BAP (μmol/L) was not significantly different between phases before (CON: 2085.9 ± 143.6, OLI: 2006.2 ± 205.5) and after (CON: 2631.9 ± 213.9, OLI: 2626.0 ± 229.7) the Wingate test and the change (%) was not different between phases (CON, 126.4 ± 9.0; OLI, 131.9 ± 15.0).
Heart rate and RPE
The maximum heart rate (bpm) after 2, 3, 4, 7, 8, 9, and 10 sets were significantly higher in the OLI phase than in the CON phase (P < 0.05, P < 0.01) (Fig. 6). Furthermore, the average heart rate (bpm) after ten sets of HIIE was also significantly higher in the OLI phase (160.3 ± 10.9) than in the CON phase (155.9 ± 10.8) (P < 0.05, d =0.40). In the Wingate test, the maximum heart rate (bpm) and the average heart rate (bpm) were not significantly different between the CON (168.1 ± 8.2 and 144.6 ± 8.5, respectively) and OLI (167.0 ± 10.9 and 147.5 ± 10.1, respectively) phases.
We observed that the RPE was not different in each HIIE set, and the mean RPE values of the ten sets did not differ between the CON (16.6 ± 1.6) and OLI (16.9 ± 1.2) phases. Furthermore, the RPE immediately after the Wingate test was not significantly different between the CON (18.9 ± 1.5) and OLI (19.3 ± 1.2) phases.
DISCUSSION
Our study revealed that seven days of OLFE supplementation positively affected HIIE performance in the trained athletes. The power output was significantly increased in the first, second, and latter sets of HIIE in the OLI phase, which suggested that short-term OLFE loading might improve HIIE performance. Although the sample size was small and it was difficult to obtain a significant difference, short-term OLFE loading resulted in a small but significant change in exercise performance. In addition, the RPE did not change between the CON and OLI phases, which suggested that the participants’ subjective exercise intensity did not increase with power output after OLFE supplementation.
With respect to energy metabolism, anaerobic metabolism was dominant in the first and second sets, and the ratio of aerobic metabolism was increased in the subsequent sets [36,40]. Therefore, OLFE loading might contribute to an increase in power output via improved anaerobic and aerobic metabolism. The heart rate was also significantly higher during HIIE in the OLI group, which indicated that the increased power output resulted in a higher heart rate. In parallel, the reduction of blood lactate after exercise in the OLI phase could be interpreted as an improvement in aerobic capacity by OLFE during HIIE.
In the present study, OLFE loading increased oxygen uptake during HIIE, as heart rate is a factor in calculating oxygen uptake according to the Fick equation [41]. Indeed, previous studies have shown that OLFE supplementation increases the maximum oxygen uptake during HIIE [32] and extends the time to exhaustion to 80% with the maximum heart rate [42]. These results suggest that OLFE supplementation can enhance aerobic capacity, and our data indirectly support this hypothesis. The mechanism of enhancing aerobic capacity by OLFE is not yet elucidated; however, some food-derived polyphenols have been shown to activate mitochondrial biogenesis and function in skeletal muscles [43,44]. Changes in blood lactate levels were significantly lower in the OLI phase despite the increased power output and heart rate during HIIE. OLFE supplementation has been shown to suppress lactate production through mitochondrial adaptation [45,46] but further studies are warranted to clarify the effects of OLFE at the molecular level.
Previous studies have shown that ROS production increases with increased exercise intensity [7-9]. Furthermore, the levels of metabolites and d-ROM were significantly elevated after high-intensity exercise compared with those at rest [47]. Here, we expected d-ROM levels to increase since power output increased after OLFE supplementation; however, the changes before and after exercise in the OLI phase were not different from those in the CON phase. These results suggest that OLFE modulates exercise-induced ROS. However, the change in BAP tended to increase after exercise with OLFE supplementation. Although there were no significant changes in BAP between the CON and OLI phases, the trend suggested that blood polyphenol concentrations increased after 7 d of supplementation with OLFE [27], reinforcing its antioxidant potential. In addition, Arent et al. [48] reported that supplementation with a food-derived polyphenol extract during HIIE with an ergometer significantly improved antioxidant properties compared to a placebo 30 and 60 min after exercise, not immediately after exercise. A significant increase in BAP was not observed because the measurement was performed only immediately after exercise in this study.
Previous studies have shown that the power output during high-intensity exercise increases upon acute [49] or 7-to 8-day [18,19] supplementation of polyphenol-rich fruits in trained males. Studies have also shown that polyphenol-rich fruits attenuate inflammation and muscle soreness during high-intensity exercise. Similarly, OLFE supplementation reduces high-intensity exercise-induced inflammation [28,29] and muscle damage [31]. Although the mechanism of action of these polyphenols remains unclear, their antioxidant potential might support training-induced inflammation.
The effects of OLFE supplementation on athletic performance have not been previously investigated. In the present study, power output during HIIE increased after seven days of supplementation with 200 mg OLFE, suggesting that short-term OLFE loading improves athletic performance. In particular, the regulation of cytosolic calcium homeostasis is a possible factor for the improvement of power output during exercise sets. Intracellular calcium concentration is regulated by calcium released from the endoplasmic reticulum in muscle cells. When the muscle is stimulated, calcium is released from the sarcoplasmic reticulum into the muscle cytoplasm through a calcium channel, the ryanodine receptor [50]. The muscle then contracts through the activation of actin-myosin interactions, and the ryanodine receptor is post-translationally modified by ROS. Since excessive ROS may prevent calcium release and attenuate muscle contraction [6], OLFE supplementation may prevent the oxidative modification of the receptor via antioxidant effects [30]. Therefore, the increase in power output in the first and second sets of HIIE in this study might be due to the antioxidant potential of OLFE, which might reduce ROS generation.
Furthermore, OLFE supplementation did not impair post-exercise performance (30-s Wingate test). It was estimated that the physical load increased according to the power output increase during HIIE, which attenuated post-exercise performance. However, this result was not expected. This result suggests that the antioxidant potential of OLFE might suppress post-exercise inflammation and promote recovery [28,29,31]. Therefore, the present study suggests that OLFE supplementation may improve HIIE performance without negative post-exercise outcomes.
This pilot study has both strengths and limitations. Its strengths include excellent adherence of participants to the intervention, with no dropouts. In contrast, there are some limitations, particularly the use of a small group of participants and the lack of a placebo group. Although the results of this open-label study are encouraging, future trials need to be designed, including a placebo group with a crossover design, to validate the effects of OLFE on high-intensity exercise performance. In addition, we were not able to conduct a dietary survey during the intervention phases, although participants were instructed to avoid supplements and antioxidants or changes to their diet.
Another limitation is that we did not measure d-ROMs and BAP during HIIE since we could not measure the changes in d-ROMs and BAP in each set during HIIE. As mentioned above, the physical exertion of the first and second sets of HIIE might be related to the antioxidant potential of OLFE. Further studies are required to measure oxidative stress and the antioxidant potential of OLFE during HIIE to clarify the relationship between OLFE intake and anaerobic metabolism. In addition, d-ROMs and BAP were the only indicators of oxidative stress and antioxidant potential. Future research should examine redox balance using other oxidative stress (e.g., carbonylated protein) and antioxidant potential (e.g., total antioxidants) markers. In addition, we could not measure the respiratory quotient (RQ) during HIIE. Therefore, we discussed anaerobic and aerobic exercise metabolism during HIIE based on previous studies. Investigation of the RQ is required to determine the change in metabolic performance upon OLFE supplementation. These investigations may accurately determine whether OLFE modulates redox balance during high-intensity exercise and promotes skeletal muscle adaptation. Despite having some limitations, our pilot study demonstrated the effects of a seven-day supplementation regimen with 200 mg OLFE on athletic performance. Hence, our study may encourage randomized, placebo-controlled, double-blind, crossover trials to assess the effects of OLFE supplementation on athletic performance in the future.
Finally, we recruited trained athletes from a university sports club who regularly exercised. Optimal OLFE intake depends on ROS production and antioxidant capacity, which are affected by individual conditioning and fitness levels. Therefore, OLFE intake should be adjusted for individuals’ age, muscle mass, and exercise habits.
In conclusion, the present study demonstrated that supplementation with a highly bioavailable polyphenol, OLFE, for seven days at 200 mg/day improves power output during HIIE without adverse effects on post-exercise performance. Therefore, HIIE performance, which requires both anaerobic and aerobic metabolism, may be improved through the antioxidant effect or activation of metabolic factors exerted by short-term OLFE loading. This strategy may contribute to the maintenance of athletic performance, especially when training intensity is increased.